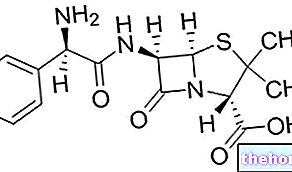
גליקוגן הוא מקרומולקולה (מסה מולקולרית של כ -400 מיליון דלטון) של גלוקוז α שבו יש בעיקר קשרי α-1,4 גליקוזידים והשלכות ביחס של 1:10, עקב קשרים α-1,6 גליקוזידים.
הגליקוגן מהווה חומר מילואים והוא מתפורר ומתחדש ללא הרף; במסת התא של כל הגוף יש כ -100 גרם של גליקוגן: רובו נמצא בכבד שבו הוא נייד ולכן יכול לשמש עתודה לאיברים אחרים (גליקוגן בשרירים אינו נייד).
האנזימים המזרזים את הפירוק והסינתזה של הגליקוגן נמצאים כולם בציטופלזמה, ולכן יש צורך במערכת ויסות שהופכת נתיב אחד לבלתי פעיל כאשר השני פעיל: אם יש גלוקוז זמין, השני הופך לגליקוגן (אנבוליזם) שהוא עתודה, להיפך, אם c "נדרש לגלוקוז, אז הגליקוגן מתפרק (קטבוליזם).

האנזים המעורב בעיקר בפירוק הגליקוגן הוא גליקוגן פוספורילאז; אנזים זה מסוגל לבקע קשר α-1,4 גליקוזידי באמצעות אורתופוספט אורגני כסוכן ליטי: המחשוף מתרחש בדרך פוספורוליטית ומתקבל גלוקוז 1-פוספט.
בחמש או שש יחידות מנקודת הסתעפות, האנזים הגליקוגן פוספורילאז אינו מסוגל יותר לפעול ולכן הוא מתנתק מהגליקוגן ומוחלף באנזים משדרש שהוא טרנספראז: באתר הקטליטי של אנזים זה c "הוא" היסטידין המאפשר העברת שלוש יחידות סוכריד לשרשרת הגליקוזידית הקרובה ביותר (היסטידין תוקף את הפחמן הראשון של מולקולת גלוקוז). האנזים שהוזכר זה עתה גליקוזיל -טרנספראז; בסוף הפעולה של אנזים זה, נשארת יחידת גלוקוז אחת בשרשרת הצדדית כאשר הפחמן הראשון קשור לפחמן השישי של גלוקוז בשרשרת הראשית. יחידת הגלוקוז האחרונה בשרשרת הצד משתחררת על ידי פעולתו של "אֶנזִים α-1,6 גליקוזידאז (אנזים זה מהווה את החלק השני של האנזים המשפיל); בהתחשב בכך שהענפים בגליקוגן נמצאים ביחס של 1:10, מההתפרקות המלאה של המקרומולקולה אנו מקבלים כ -90% מהגלוקוז 1-פוספט וכ -10% של גלוקוז.
פעולתם של האנזימים הנ"ל מאפשרת חיסול של שרשרת צדדית ממולקולת הגליקוגן; ניתן לחזור על פעילותם של אנזימים אלה עד להתרבות מוחלטת של השרשרת.
בואו ניקח בחשבון הפטוציט; גלוקוז (נטמע באמצעות התזונה), כאשר הוא נכנס לתא הופך לגלוקוז 6-פוספט ובכך מופעל. גלוקוז 6-פוספט, על ידי פעולה של פוספוגוקומוטאז, הופך לגלוקוז 1-פוספט: האחרון הוא מבשר לא מיידי של ביוסינתזה; בביוסינתזה משמשת צורה מופעלת של סוכרים המיוצגת על ידי סוכר המקושר לדיפוספט: בדרך כלל uridyldiphosphate (UDP). גלוקוז 1- פוספט הוא לאחר מכן הופך לגלוקוז UDP, מטבוליט זה תחת פעולה של גליקוגן סינתז אשר מסוגל לקשור UDP-גלוקוז לקצה בלתי-מצמצם של הגליקוגן הגדל: מתקבלים גליקוגן מוארך של יחידה גלוקוזידית ו- UDP. UDP הופך על ידי האנזים הדיפוספוקינאז הנוקלאוצידי ל- UTP שחוזר למחזור הדם.
התדרדרות הגליקוגן מתרחשת על ידי הפעולה של גליקוגן פוספורילאז המשחרר מולקולה של גלוקוז והופך אותה לגלוקוז 1-פוספט. לאחר מכן, phosphoglucomutase ממיר גלוקוז 1-פוספט לגלוקוז 6-פוספט.
הגליקוגן מסונתז, בעיקר, בכבד ובשרירים: באורגניזם יש 1-1.2 ק"ג גליקוגן המופץ לאורך מסת השריר.
הגליקוגן של מיוציט מייצג עתודת אנרגיה רק לתא זה בעוד הגליקוגן הכלול בכבד הוא גם עתודה לרקמות אחרות, כלומר ניתן לשלוח אותו, כגלוקוז, לתאים אחרים.
גלוקוז 6-פוספט המתקבל בשרירים מפירוק הגליקוגן נשלח לאחר מכן, במקרה של דרישת אנרגיה, לגליקוליזה; בכבד, גלוקוז 6-פוספט הופך לגלוקוז על ידי פעולה של גלוקוז 6-פוספט פוספטאז (אנזים אופייני של הפטוציטים) ומועבר למחזור הדם.
גליקוגן סינתאז וגליקוגן פוספורילאז פועלים שניהם על יחידות הגליקוגן שאינן מפחיתות ולכן חייב להיות אות הורמונלי המצווה על הפעלת מסלול אחד וחסימתו של השני (או להיפך).
במעבדה ניתן היה להאריך את שרשרת הגליקוגן על ידי ניצול גליקוגן פוספורילאז ושימוש בגלוקוז 1-פוספט בריכוז גבוה מאוד.
בתאים, זרחן הגליקוגן זרז רק את תגובת הפירוק מכיוון שריכוזי המטבוליטים הם כך שיזנו את שיווי המשקל של התגובה הבאה ימינה (כלומר לעבר פירוק הגליקוגן):

בואו נראה את מנגנון הפעולה של הפוספורילאז הגליקוגן: החמצן האצטלי (הפועל כגשר בין יחידות הגלוקוז) נקשר למימן של הפוספוריל: נוצר ביניים לתגובה הניתנת על ידי פחמימות (על הגלוקוז שהוא כולו " הגפיים) שאליהן נקשר הזרחון (Pi) במהירות רבה.
זרחן גליקוגן דורש קופקטור שהוא פוספט פירידוקסלי (מולקולה זו היא גם קופקטור לטרנסמינאזות): יש לה פוספוריל מוגן חלקי (פוספט פירידוקס מוקף בסביבה הידרופובית המצדיקה את נוכחותם של פרוטונים הקשורים אליו). הפוספוריל (Pi) מסוגל להעביר פרוטון לגליקוגן מכיוון שפוספוריל זה רוכש מחדש את הפרוטון מהזרחן הפרוטוני המפוגן חלקית של הפוספט הפירידוקסלי. ההסתברות שב- pH פיזיולוגי הפוספוריל מאבד את הפרוטון שלו ונותר מופחת לחלוטין הוא נמוך מאוד.
בואו נראה כעת כיצד פועל phosphoglucomutase.אנזים זה מציג, באתר הקטליטי, שאריות של סרין זרחני; סרין מניב פוספוריל לגלוקוז 1-פוספט (במיקום שש): גלוקוז 1,6-ביספוספט נוצר לזמן קצר, ואז הסרין מתחדש ולוקח את הפוספוריל במיקום הראשון. מוטאז פוספוגלוקו יכול לפעול לשני הכיוונים, כלומר להמיר גלוקוז 1-פוספט לגלוקוז 6-פוספט או להיפך; אם נוצר גלוקוז 6-פוספט, ניתן לשלוח אותו ישירות לגליקוליזה, בשרירים, או להפוך לגלוקוז בכבד.
האנזים transidase uridyl phosphogluco (או pyrophosphorylase גלוקוז UDP) מזרז את תגובת העברת הגלוקוז 1-פוספט ל- UTP על ידי הצמדה לפוספוריל א.
האנזים שתואר זה עתה הוא פירופוספורילאז: שם זה נובע מכך שהתגובה ההפוכה לאחת שתוארה זה עתה היא פירופוספורילציה.
גלוקוז UDP, המתקבל כמתואר, מסוגל להאריך את שרשרת הגליקוגן, על ידי יחידה חד -סוכרית.
אפשר לגרום לתגובה להתפתח לקראת יצירת גלוקוז UDP על ידי חיסול מוצר שהוא פירופוספט; האנזים פירופוספטאז הופך את הפירופוספט לשתי מולקולות של אורתופוספט (הידרוליזה של אנהידריד) ובכך שומר על ריכוז הפירופוספט נמוך כל כך עד שתהליך היווצרותו של גלוקוז UDP מועדף תרמודינמית.
כאמור, גלוקוז UDP, הודות לפעולה של גליקוגן סינתז, מסוגל להאריך את שרשרת הגליקוגן.
ההשלכות (ביחס של 1:10) נובעות מכך שכאשר שרשרת גליקוגן מורכבת מ- 20-25 יחידות, אנזים מסתעף (בעל "היסטידין באתר הקטליטי שלו) מתערב, המסוגל להעביר סדרה של 7 -8 יחידות גליקוזידיות יותר במורד הזרם של 5-6 יחידות: כך נוצרת הסתעפות חדשה.
מסיבות של מוצא עצבי או אם יש צורך באנרגיה עקב מאמץ גופני, אדרנלין מופרש מבלוטת יותרת הכליה.
תאי המטרה של אדרנלין (ונוראדרנלין) הם תאי הכבד, השרירים ורקמת השומן (באחרונה יש התדרדרות של טריגליצרידים ומחזור הדם של חומצות שומן: כתוצאה מכך, גלוקוז מיוצר במיטוכונדריה 6 -פוספט, שיהיה נשלח לגליקוליזה, בעוד שבאדיפוציטים גלוקוז 6-פוספט הופך לגלוקוז על ידי פעולת האנזים גלוקוז 6-פוספט פוספטאז ומיוצא לרקמות).
בואו לראות, עכשיו את שיטות הפעולה של האדרנלין. אדרנלין נקשר לקולטן הממוקם על קרום התא (של מיוציטים והפטוציטים) וזה קובע את תרגום האות מבחוץ לתא. מופעל חלבון הקינאז הפועל בו זמנית על המערכות המסדירות סינתזה ופירוק הגליקוגן:
הגליקוגן סינתז קיים בשתי צורות: צורה דהפוספורילאלית (פעילה) וצורה זרחנית (לא פעילה); חלבון קינאז זרחן גליקוגן סינתאז וחוסם את פעולתו.
גליקוגן פוספורילאז יכול להתקיים בשתי צורות: צורה פעילה בה נמצא סרין זרחני וצורה לא פעילה בה הסרין מתפענח. ניתן להפעיל את הגליקוגן פוספורילאז על ידי האנזים גליקוגן פוספורילאז קינאז. הגליקוגן פוספורילאז קינאז פעיל אם הוא פוספורילציה ולא פעיל אם הוא מפוספיריל; לחלבון קינאז יש כמצע את הגליקוגן פוספורילאז קינאז, כלומר הוא מסוגל לפוספורילט (ולפיכך להפעיל) את זה אשר בתורו מפעיל את הפוספורליאז הגליקוגן.
ברגע שאות האדרנלין נגמר, ההשפעה שיש לו על התא חייבת להסתיים גם היא: אנזימי פוספטאז מתערבים אז על מיני החלבונים.